55. Did Man Migrate Out of Africa and All Over the Earth Starting 60,000 Years Ago?
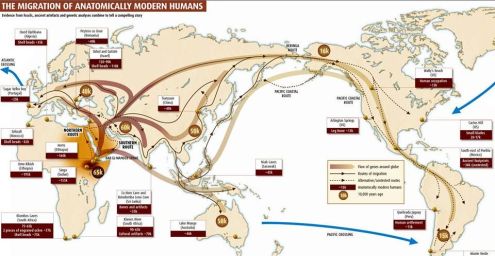 Above is what I call a “map with arrows” that purportedly shows how manking migrated all over the earth 60,000 years ago from central Africa. Maps with arrows are incredibly easy to create. The path these kinds of maps show are impossible. Humans did not and could not have migrated out of Africa 60,000 years ago to populate the entire globe with billions of people as NatGeo, the Smithsonian, Scientific American, every anthropology class and textbook, and virtually every scientist on earth says they did. The journey, no matter how long it took, would have killed all of the trekkers before they conquered the first major obstacle, the Sahara Desert. My video documentary uses a NatGeo documentary, The Journey of Man: A Genetic Odyssey, to make its points. You can see the whole NatGeo documentary below my video.:
Above is what I call a “map with arrows” that purportedly shows how manking migrated all over the earth 60,000 years ago from central Africa. Maps with arrows are incredibly easy to create. The path these kinds of maps show are impossible. Humans did not and could not have migrated out of Africa 60,000 years ago to populate the entire globe with billions of people as NatGeo, the Smithsonian, Scientific American, every anthropology class and textbook, and virtually every scientist on earth says they did. The journey, no matter how long it took, would have killed all of the trekkers before they conquered the first major obstacle, the Sahara Desert. My video documentary uses a NatGeo documentary, The Journey of Man: A Genetic Odyssey, to make its points. You can see the whole NatGeo documentary below my video.:

You must be logged in to post a comment.